
Строение мышечных волокон и механизм мышечных сокращений.
Статическая сила, динамическая сила, статическая силовая выносливость, динамическая силовая выносливость… — физические качества, уровень развития которых определяет спортивный результат в подтягивании.
Миофибриллы, митохондрии, саркоплазматический ретикулум… — структурные элементы мышечной клетки, участвующие в преобразовании потенциальной химической энергии в полезную механическую работу или мышечное напряжение.
Креатинфосфатная реакция, гликолиз, аэробное окисление – механизмы энергообеспечения, которые служат делу обеспечения непрерывного ресинтеза АТФ в работающих мышцах.
Медленные окислительные, быстрые гликолитические, быстрые окислительно-гликолитические – типы мышечных волокон, отличающихся по скорости сокращения, активности ферментов ресинтеза АТФ, преимущественным механизмам энергопродукции.
Попробуем увязать между собой физические качества спортсмена, физиологию мышечного сокращения и биохимические процессы, происходящие в мышечных клетках. Для этого предварительно рассмотрим строение мышечного волокна и механизм мышечного сокращения в той степени, в которой это необходимо для подтягиваний.
01.1_ Строение и химический состав скелетных мышц
Скелетная мышца состоит из мышечных волокон (миоцитов). Мышечные волокна представляют собой гигантские многоядерные клетки длиной от 0,1 до 2-3 сантиметров, а в некоторых мышцах миоциты достигают 12 сантиметров. Площадь поперечного сечения мышечных клеток составляет от 3 до 10 квадратных микрометров.
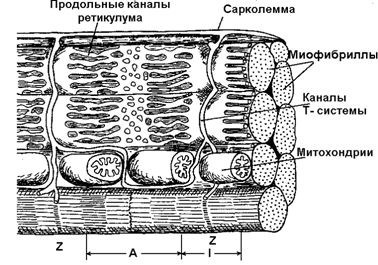
Волокно покрыто эластичной оболочкой — сарколеммой и состоит из саркоплазмы, структурными элементами которой являются такие органоиды, как митохондрии, рибосомы, трубочки и пузырьки саркоплазматической сети (ретикулума) и так называемая Т-система а также различные включения. В саркоплазме условно выделяют две части – саркоплазматический матрикс и саркоплазматический ретикулум. Саркоплазматический ретикулум, представляющий собой определённым образом организованную сеть соединяющихся цистерн (содержащих в большой концентрации ионы кальция) и трубочек, играет важную роль в механизмах сокращения и расслабления мышцы. Кроме того, к части ретикулума прикреплены рибосомы, специальные сферические образования, на которых и при участии которых происходит биосинтез белков.

Саркоплазматическая сеть с помощью особых трубочек, называемых Т-системой, связана с оболочкой мышечной клетки. Т-система также имеет прямое отношение к мышечному сокращению, так как по ней передаётся изменение электрического потенциала поверхностной мембраны элементам ретикулума, что приводит к освобождению ионов кальция, поступающих к миофибриллам и запускающих процесс мышечного сокращения.

Рисунок 1. Строение Т-системы и саркоплазматического ретикулума мышечного волокна [по: Кроленко, 1975].
Саркоплазматический матрикс представляет собой коллоидный раствор, содержащий белки, гликоген, жировые капли и другие включения. Миофибриллы – сократительные элементы мышечных клеток – также находятся в саркоплазматическом матриксе.
Кроме того, в саркоплазме находятся ферменты гликолиза, расщепляющие гликоген или глюкозу до пировиноградной или молочной кислоты и креатинкиназа – фермент, ускоряющий креатинфосфатную реакцию. Особый белок саркоплазмы – миоглобин – обеспечивает некоторый запас кислорода в мышечной ткани, а также участвует в переносе кислорода от сарколеммы к митохондриям.
Мышечная клетка имеет не одно, а множество ядер, которые располагаются на её периферии – под сарколеммой. Внутри каждого ядра находится ДНК, являющаяся носителем носледственной информации и состоящая из генов, в которых закодирована структура всех синтезируемых мышечными волокнами белков.
Лизосомы, представляющие собой микроскопические пузырьки, содержат в растворённом виде различные ферменты, способные в условиях кислой реакции среды расщеплять различные высокомолекулярные вещества. Такая необходимость может возникать в мышечных клетках, например, при очень напряжённой мышечной деятельности.
01.2_Митохондрии
Митохондрии, одни из важнейших структурных компонентов мышечного волокна, располагаются цепочками вдоль миофибрилл (рисунок 1.), тесно соприкасаясь с мембранами ретикулума. В митохондриях протекает аэробное окисление углеводов, жиров и аминокислот, а за счёт энергии, выделяющейся при окислении, происходит ресинтез АТФ.
Митохондрии ограничены двумя мембранами (рисунок 2.). Наружная митохондриальная мембрана имеет ровные контуры, не образует выпячиваний или складок. Наружную мембрану от внутренней отделяет межмембранное пространство. Внутренняя мембрана ограничивает внутреннее содержимое митохондрии, ее матрикс. Характерной чертой внутренней мембраны митохондрий является их способность образовывать многочисленные выпячивания внутрь митохондрий. Такие выпячивания чаще всего имеют вид плоских гребней, или крист, существенно увеличивая поверхность внутренней мембраны. Мембраны митохондрий построены из белка и содержащих фосфорную кислоту жироподобных веществ — фосфолипидов. На внутренней мембране в определённом порядке расположены биологические катализаторы – ферменты, при помощи которых происходят окислительные процессы, а также компоненты дыхательной цепи – главной системы превращения энергии в митохондриях. На внешней мембране митохондрий в определённом порядке расположены ферменты, не имеющие отношения к дыхательной цепи. Немало ферментов в растворённом виде содержится и в матриксе. Кроме того, матрикс митохондрий содержит рибосомы и митохондриальную ДНК.

о А.Кузнецов)
Долгое время было непонятно, почему так происходит. В качестве одной из возможных причин называлась перегрузка нервной системы тренировками до отказа. Но срыв адаптации обычно происходил на фоне эмоционального подъёма о